2023年11月20日
茨城工業高等専門学校
京都大学複合原子力科学研究所
日本原子力研究開発機構
茨城大学
タンパク質の分子骨格が実は持っていた知られざる機能
中性子で明らかに
タンパク質分子の機能の解明や、迅速な創薬に向けた新しいタンパク質分子デザインに期待
【発表のポイント】
- 日本原子力研究所の生体高分子用単結晶中性子回折装置-3(BIX-3)を用いて、ヒトリゾチームの中性子結晶構造解析(※1)実験を行ない、タンパク質分子の骨格に重大な影響を及ぼす水素を発見しました。
- 中性子で観測された水素イオンや重水素イオンにより、長年、一様な平面として扱われてきたペプチド結合(※2)の平面性が個々に緩和されることを見出しました。
- 核磁気共鳴(NMR)法(※3)で観測した15Nのケミカルシフトデータから、平面性が失われたペプチド結合では、電子共鳴構造が壊れていることが示されました。
- 中性子で新たな水素が観測された部位では、タンパク質分子自身が作った負の静電ポテンシャル(※4)により、ペプチド結合の酸素原子と水素の相互作用が強められたと考えられます。
- 全てのタンパク質分子で利用されうるペプチド結合の性質の変化の発見は、創薬にむけたタンパク質分子デザイン法の革新の礎になると期待されます。
茨城工業高等専門学校教授 千葉薫(ちばかおり)、元日本原子力研究所(現、日本原子力研究開発機構原子力科学研究所)特別研究員 松井拓郎(まついたくろう)、京都大学複合原子力科学研究所准教授 茶竹俊行(ちゃたけとしゆき)、日本原子力研究開発機構 元量子ビーム応用研究部門(現、J-PARCセンター)研究主幹 大原高志(おおはらたかし)、故、理化学研究所上級研究員 油谷克英(ゆたにかつひで)、茨城大学教授 田中伊知朗(たなかいちろう)、元日本原子力研究所研究主幹、元茨城大学教授 新村信雄(にいむらのぶお)は、日本原子力研究所(現、日本原子力研究開発機構原子力科学研究所)の生体高分子用単結晶中性子回折装置-3(BIX-3)を用いて、ヒトリゾチーム(130アミノ酸からなる分子量14.3kDaのタンパク質分子)の中性子結晶構造解析実験を行ないました。
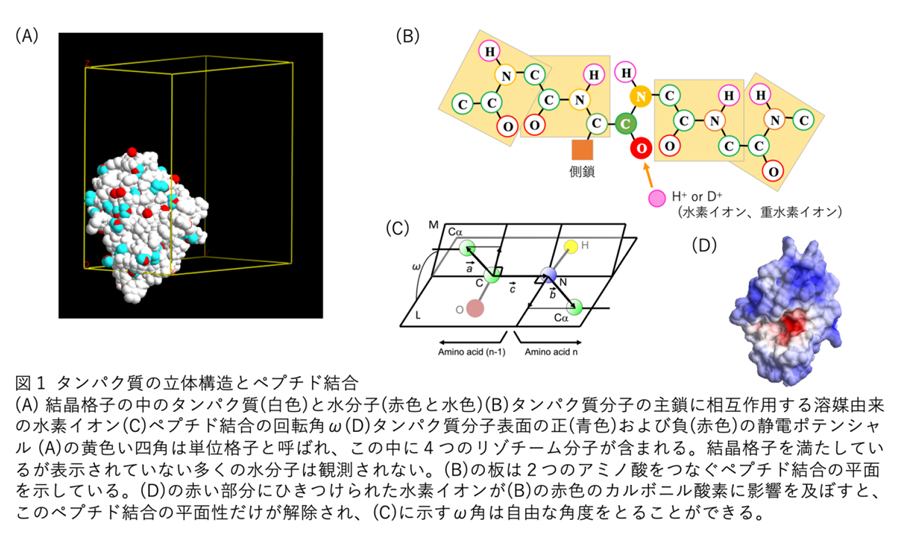
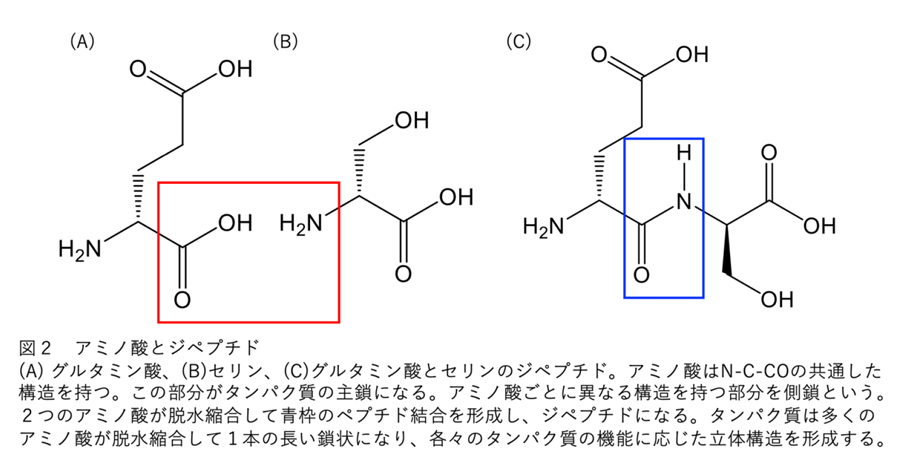
タンパク質はアミノ酸が1本の鎖状につながった巨大分子です(図1A)。これまで、タンパク質分子の骨格である主鎖(※5)を支えるペプチド結合は、Paulingらが調べた小さな有機化合物の中のペプチド結合(図2)と同じく、1951年以降現在まで、一様な硬さの平面構造をとると考えられてきました。プロテインデータバンク(PDB)に登録された20万個以上のタンパク質分子の立体構造解析も、原則的にはペプチド結合が平面であるという前提で行われています。しかし、本研究で行なったヒトリゾチームの中性子結晶構造解析を起点として、複数の実験手法で得られたデータを精査したところ、タンパク質の分子の中では、タンパク質分子自身が作る静電ポテンシャル(図1D、※4)などにより作られる「タンパク質場」に引き寄せられた水素イオンまたは重水素イオンによって、ペプチド結合が個々に柔らかくなることがわかりました。本研究の結果は、タンパク質のペプチド結合は一様に硬い平面である、という、長年にわたりタンパク質の構造研究を支えてきた概念と、タンパク質の主鎖は立体構造の骨格を保ち、側鎖(※5)が機能を担う、という役割分担を見直す必要があることを示しています。
本研究成果は、アメリカ蛋白質学会(The Protein Society)発行の『Protein Science』に”Site-specific relaxation of peptide bond planarity induced by electrically attracted proton/deuteron observed by neutron crystallography”として9/20(水)<現地時間>にオンラインで掲載されました。
(1) これまでの研究でわかっていたこと
タンパク質は、小さいものでも1000個以上の原子からなる巨大分子です。タンパク質の構造解析では、タンパク質分子を形作る各々の原子の位置(座標)を求めます。よく使われる方法は、タンパク質水溶液を使って作製したタンパク質の結晶にX線をあて、得られた回折データから原子座標を計算で求めるX線結晶構造解析(※1)です。X線のかわりに中性子を使えば、中性子結晶構造解析になります。このほかに、核磁気共鳴(NMR)法や電子顕微鏡を使ってタンパク質の構造解析を行う方法もあります。タンパク質の立体構造データを集めたプロテインデータバンク(PDB)に最も多くエントリーされているのはX線結晶構造解析で求めた原子座標データです。X線は電子で散乱するため、X線結晶構造解析では電子を持たない水素イオンや重水素イオンは観測できませんが、中性子は原子核で散乱するため、X線では観測できない水素イオンや重水素イオンの原子核の位置を観測することができます。この特長を生かして中性子結晶構造解析は、主にタンパク質の働きに重要な役割を果たす側鎖の水素を観測する実験に用いられてきました。
タンパク質の結晶構造解析も、小さな分子の結晶構造解析と同じ原理を使っていますが、原子の数が多すぎるため、実験データを解析する際、既にわかっている基本情報を与えて計算を行う点が異なります。例えば、実験試料となるタンパク質の中で、どんな形のアミノ酸がどういう順番につながっているかという情報や、C-H、C-Nなどの原子間結合距離や結合角などの情報です。「ペプチド結合は平面である」という情報もタンパク質の構造解析には必要不可欠な情報です。例外的にとても分解能の高いデータが手に入り、このペプチド結合は曲げた方がデータとよく合う、ということがわかったペプチド結合に限り、曲がってもよいとする場合がほとんどです。結晶構造解析では、たくさんの分子の平均の構造を観測するので、分子ごとにあちこちを向いている側鎖などは、平均の構造では1つの座標に決めることができません。多くの分子でこちらを向いている、ということが観測された場合のみ、そちらの方向に原子座標を置きます。タンパク質の主鎖は側鎖に比べて動きにくいので、どの分子でも同じ位置にあることが多く、結晶構造解析で比較的観測しやすいですが、主鎖の構造もはっきりわからない部分のペプチド結合は平面のままにしておくのが普通です。近年、高分解能の構造解析が行えるようになってきたため、構造解析で得られたタンパク質の分子モデルにもしばしば歪んだペプチド結合が観測されるようになってきました。しかし、これらのペプチド結合がなぜ歪んでいるか、その化学的な原因についてはわかっていませんでした。
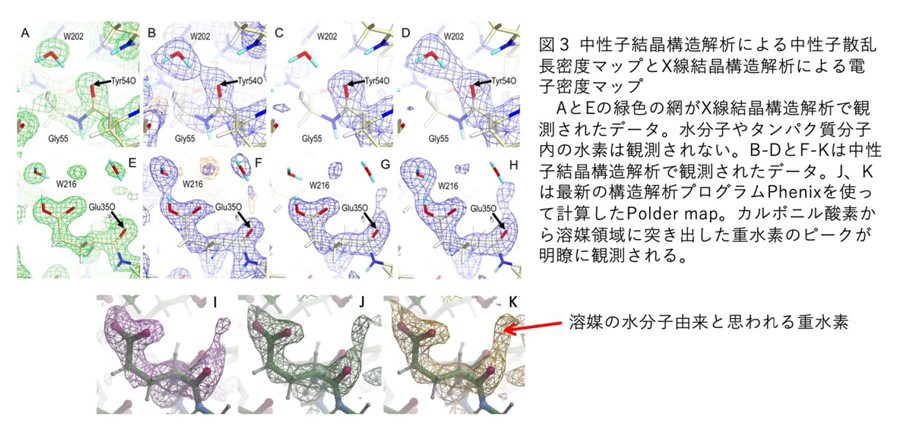
結晶中に並んだタンパク質分子のすき間を満たす多くの水分子のうち、結晶構造解析で原子座標が決められるのは、タンパク質分子の中のある原子にひきつけられ、同じ位置に留まりやすくなったわずかな水分子だけです(図3)。さらに、X線結晶構造解析では水素はほぼ観測できないため、酸素原子がタンパク質分子にひきつけられるなどして、どの結晶格子の中でも同じ位置にあるわずかな水分子(図3A、E)以外は観測されません。タンパク質の結晶の中には、このような観測できない水分子が数多く含まれています(図1Aの空白部分にも水分子が満たされています)。水素を観測することができる中性子結晶構造解析では、タンパク質分子の酸素に水素がひきつけられた水分子の、水素だけが観測される場合があります(図3F-K)。中性子結晶構造解析の際、このような酸素の位置が定まらない水分子をどう扱うか、最も慎重に判断しながら解析を進める必要があります。
NMR法も結晶構造解析とならんで、タンパク質分子中の原子を個別に観測することができる有力な手法です。本研究で中心的な役割を果たす主鎖アミド窒素の15Nケミカルシフトは、タンパク質の分子内環境の変化を敏感に反映するといわれていますが、化学的などのような変化を反映しているかはわかっていませんでした。
(2) 今回の研究であらたに実現しようとしたこと、明らかになったこと
本研究では、ヒトリゾチーム分子の中性子結晶構造解析を行い、結晶中の個々の分子ごとに構造が変わりにくい主鎖に注目して、中性子結晶構造解析で得られた中性子散乱長密度マップ(※1)を慎重に調査しました。その結果、ペプチド結合をつくるカルボニル基の酸素の近くに、重水素らしきピークを発見しました(図3F-K)。タンパク質分子の結晶は、位置が定まらない水分子で満たされていて、これらをどのように計算に入れるかによってマップの形が少しずつ変わります。図3F-Kのどの図でも、右側の酸素のところから伸びているピークが見えますが、Phenixという最新の構造解析プログラムでPolder Mapという図を描くと、さらにはっきりとこの重水素が描き出されることがわかります。おそらく水分子の水素が、タンパク質の酸素原子にひかれてこの位置に留まりやすくなっているのに水分子の酸素を引きつける原子が近くになかったため、水分子の酸素原子の位置が定まらず、水素だけが観測されたのであろうと考えました。タンパク質の結晶構造解析でこのような重水素を観測したのも、主鎖近傍の重水素に注目して研究を進めたのも私たちが世界ではじめてです。
(3) そのために新しく開発した手法
本研究では中性子結晶構造解析データの他に、NMRやX線結晶構造解析など、他の実験で得られたデータや、タンパク質の原子座標から計算した静電ポテンシャルを用いています。新しく発見されたこの水素が、タンパク質分子の構造だけでなく、タンパク質分子をつくる化学結合にどのような影響を及ぼしているか、また、なぜこの位置に水素イオンが溜まりやすくなっているかを総合的に考えたところに特徴があります。
タンパク質の結晶構造解析では、マップを描く時の条件によってピークの形が変わることからもわかるように、図3の中性子のピークを観測しただけでは、「ここに重水素がある」と断言することはできません。そこで私たちはまず、「ここに水素があったら何が起こるか」を考えました。
最初に着目したのはNMR法です。なかでも、窒素のNMRデータに注目しました。全てのアミノ酸は、ペプチド結合でつながってタンパク質になった時に、主鎖になる部分に1つずつ窒素原子を持っています(図2)。2つのアミノ酸がつながってペプチド結合をつくると、この窒素原子の電子の一部が結合に使われ、電子共鳴構造を作ります。そうすると、窒素原子核のまわりの電子が、ペプチド結合する前に比べて薄くなり、その結果、ペプチド結合をしている窒素の「ケミカルシフト」という値が大きくなる(この状態を低磁場シフトしている、という)ことがわかっています。タンパク質の主鎖の窒素は、一番尻尾のアミノ酸を除いて、全てペプチド結合しており、理想的には全ての窒素の「ケミカルシフト」が低磁場シフトしているはずです。本研究で観察された重水素は、タンパク質の酸素原子とある程度強い相互作用をしていると考えられるため(図1B)、この重水素がペプチド結合の電子共鳴構造を壊したとすると、この窒素の「ケミカルシフト」は相対的に高磁場シフトすると考えられます。ヒトリゾチームのGlu35とペプチド結合しているSer36の窒素のケミカルシフトデータを調べてみたところ、pH3.8からpH5.0のどのデータでも、高磁場シフトしていることがわかり、Glu35とSer36をつなぐペプチド結合の電子共鳴構造が壊れていることが示唆されました。
次に、ペプチド結合の電子共鳴構造が壊れていたら何が起こるかを考えてみました。そもそも、ペプチド結合が平面構造をしているのは、C=OとNの間に電子共鳴構造ができているからです。逆にいうと、電子共鳴構造が壊れているペプチド結合は平面でなく、図1Cのωはあらゆる角度を自由にとることができます。そこで、私たちの中性子結晶構造解析と似通った実験条件で得られた、分解能の高いX線結晶構造解析データを慎重に解析したデータを調べたところ、このペプチド結合が明らかに曲がっていることがわかりました。X線結晶構造解析のデータでペプチド結合が明らかに曲がっていたその他の10箇所のペプチド結合の酸素にも、中性子結晶構造解析で水素や重水素が観測されていることがわかりました。これらの水素は水分子の一部として観測されたり、タンパク質に結合した水素として観測されたりしていましたが、どの水素もペプチド結合を自由回転させる効果をもつと考えられます。
ペプチド結合が柔らかくなる現象は、タンパク質分子の中の全てのペプチド結合で起こるわけではありません。私たちは最後に、この現象がどのような条件が揃うと起こるのかを考えてみました。今回中性子結晶構造解析で観測された水素は、他の水素より少しだけ強く、ペプチド結合している酸素原子と相互作用していると考えられます。リゾチームの立体構造から、分子内の静電ポテンシャルを計算してみたところ、このような水素が観測される部位は負の静電ポテンシャルを示すことがわかりました。これらの結果から、ペプチド結合の酸素原子周辺に作られた負の静電ポテンシャルが、正電荷をもつ水素イオンをひきつけ、このペプチド結合だけに化学的な影響を及ぼす原因だと考えられます。
(4) 研究の波及効果や社会的影響
本研究により、中性子結晶構造解析で観測された水素と、タンパク質分子自身がつくる静電ポテンシャル、そして15NのNMRのケミカルシフトはペプチド結合の柔らかさと密接な関係があることがわかってきました。ペプチド結合の平面性は長年にわたりタンパク質の構造研究を支えてきた基本概念ですが、タンパク質の分子内静電ポテンシャルを計算するだけで予測できるペプチド結合の化学的性質の変化をとりいれることで、超高分解能データを得ることができないタンパク質でも、その構造や性質をより正しく理解し、利用することができるようになると考えられます。また、タンパク質の分子骨格の柔らかさをペプチド結合ごとに調節することで、小さな薬剤分子により強く、より選択的に結合できるタンパク質分子デザインも可能になります。本研究の結果は、迅速な新薬開発にむけたタンパク質の新たな分子デザインにも大いに役立つと期待されます。
(5) 今後の課題
本研究で観測された重水素のように、タンパク質分子を取り囲む水分子の水素がタンパク質の分子骨格に作用するという現象は、全てのタンパク質で起こっていると考えられます。現在、タンパク質の分子デザインや分子シミュレーションで「水和水」として扱っている個々の水分子が、タンパク質分子とどのような相互作用をするのか、実験と計算科学の両側からアプローチを進める必要があります。さまざまな特徴を持つタンパク質分子で、これらの相互作用がどのように利用されているかを個々に解明することで、水中でのタンパク質分子がもつ性質への理解がさらに深まると考えられます。
(6) 研究者のコメント
中性子結晶構造解析は水素を観測できる有用な実験方法です。これまでのタンパク質中性子結晶構造解析では、結晶構造解析で観測しにくい動きやすい側鎖の水素に着目した研究が主流でした。本研究では、研究対象を動きにくい主鎖に絞り、NMRなど、結晶構造解析以外の手法を、その測定原理に遡って組み合わせて用いることにしました。その結果、全てのタンパク質で利用できる、タンパク質の主鎖が実は持っていた知られざる機能を見出すことができました。今後は、ペプチド結合が柔らかくなるという現象が、生命活動の中でどのように利用されているかを探究するとともに、この原理を生かしたタンパク質分子デザインの実現にチャレンジしていきたいと考えています。
(7) 用語解説
※1【中性子結晶構造解析とX線結晶構造解析】
タンパク質の結晶に中性子をあてると、タンパク質分子の各原子によって散乱された中性子により回折データが得られます。これを解析すると、重水素(D)は窒素(N)、酸素(O)、炭素(C)と同様に正のピークとして観測されます。これに対して軽水素(H)は負のピークとして観測されます。Hはこのほか「非干渉性散乱」という、データのノイズを増やす作用ももつため、中性子結晶構造解析ではできるだけHをDに置き換えたタンパク質溶液を用意して作った結晶で実験を行います。本研究で得られた中性子結晶構造解析で溶媒の水がD2Oとして観測されるのはこのためです。タンパク質分子中にも、水分子の水素とたえず交換し続けている水素があり、これらの水素もDとして観測されます。X線結晶構造解析では、X線が電子で散乱されるため、実験データから得られる電子密度マップ(図3A、E)では電子を持たない水素イオン(H+:陽子)は観測されません。この電子密度マップに対して、中性子結晶構造解析データから得られるマップは中性子散乱長密度マップと呼ばれます(図3B-D、F-K)。電子密度マップと比較すると、側鎖の酸素に結合した水素や水分子の水素も非水素原子と同様に観測されていることがわかります。
※2【タンパク質とペプチド結合】
タンパク質は多数のアミノ酸が枝分かれのない1本の鎖状につながった巨大分子です。隣同士のアミノ酸は、各々のアミノ酸がもつカルボキシ基とアミノ基が脱水縮合した「ペプチド結合」でつながっています。Paulingらは2つのアミノ酸からなるジペプチドなどを用いた有機低分子X線結晶構造解析を行い、カルボニル基のπ電子と、アミド窒素の孤立電子対が電子を出し合った電子共鳴構造を作るため、平面構造をとることを明らかにしました。巨大分子であるタンパク質の結晶構造解析では、現在でもこの低分子有機化合物を使った構造解析結果に基づき、「ペプチド結合は平面である」という束縛のもとで分子モデルを組み立てます。近年では、分解能の高い結晶構造解析が可能となってきたため、平面でない歪んだペプチド結合の方が実験データとよく合う部位についてはこの歪みをとりいれた分子モデルがデータベースにも多く見られるようになりました。
※3【核磁気共鳴(NMR)法】
NMR法もタンパク質分子の中の水素を個々に観測することができる重要な実験方法ですが、本研究ではペプチド結合に含まれる窒素のケミカルシフトを利用しました。タンパク質のNMRを測定する際には、分子中のNを15Nに置き換えた分子を作製して実験を行います。タンパク質の15Nのケミカルシフトは、分子内部の環境によって大きく変わることが知られています。
※4【静電ポテンシャル】
アミノ酸の中には正電荷や負電荷をもつものがあります。タンパク質分子は1列につながったアミノ酸が水中で折り畳まれて立体構造を形成するため、できあがったタンパク質分子の中で正電荷や負電荷が密集するところができます。正電荷が密集するところは静電ポテンシャル(電位)が高く、負電荷が密集するところは静電ポテンシャルが低くなります。タンパク質表面の静電ポテンシャルのむらは、タンパク質が薬剤など、他の分子と結合するときに重要な目印になることがわかっています。本研究ではこの静電ポテンシャルがペプチド結合の柔らかさの調節に使われていることがわかりました。
※5【タンパク質の主鎖と側鎖】
全てのアミノ酸はアミノ基とカルボキシ基を持っています。これらがペプチド結合でつながることで、タンパク質分子は…N-Cα-(CO)-N-Cα-(CO)…という繰り返し配列の骨格構造をもつことになります。これをタンパク質の主鎖といいます。この中で(CO)とNをつなぐのがペプチド結合です。これに対してCα-(CO)とN-Cαの結合は自由回転することができます。一方、Cα原子には、各々のアミノ酸がもつ性質の異なるユニットが結合しています。これを側鎖とよび、タンパク質分子の機能は、主に側鎖を用いて説明されてきました。
(8) 論文情報
雑誌名:Protein Science
論文名:Site-specific relaxation of peptide bond planarity induced by electrically attracted proton/deuteron observed by neutron crystallography
掲載日時(現地時間):2023年9月20日
掲載URL:https://doi.org/10.1002/pro.4765
DOI:10.1002/pro.4765
(9) 研究助成
研究費名:日本原子力研究所(現、日本原子力研究開発機構)黎明研究
研究課題名:中性子結晶構造解析による蛋白質分子内静電ポテンシャルの実測
研究代表者名:千葉(鴨志田) 薫
研究費名:科学技術振興調整費開放的融合研究推進事業
研究課題名:水素・水和構造を含めた新しい構造生物学の開拓
研究代表者名:新村 信雄